D. Cillo, C. Ancona, L. Dessi, J. Matějíček, E. Bazzato
RIASSUNTO
Vengono forniti i dati relativi alle specie di Hydroadephaga presenti nel territorio Montano di Dolianova (Provincia di Cagliari, Sardegna Orientale).
ABSTRACT
Data are provided on the Hydroadephaga species present in the mountain territory of Dolianova, (Province of Cagliari, Eastern Sardinia).
Parole chiave: Coleoptera, Hydroadephaga, Italia, Sardegna, Dolianova.
Key words: Coleoptera, Hydroadephaga, Italy, Sardinia, Dolianova.
INTRODUZIONE
Nell’arco del secolo scorso (sin da Kolkwitz & Marsson, 1909) si è compreso come per determinare lo stato di salute di un corso d’acqua occorresse guardare con attenzione alle comunità animali che lo popolavano. Il concetto è stato poi esteso anche agli altri habitat e a tali animali, o comunità di animali, si è dato il nome di “Bioindicatori”. Gli invertebrati si sono rivelati a tale scopo soggetti ideali e gli Insetti si sono ritrovati ad essere considerati fra gli indicatori ambientali più importanti. Con gli anni si sono affinati metodi e differenti indici, tra i quali il più noto è rappresentato dall’Indice Biotico Esteso (IBE), che vengono (D. lgs. 152/99) usualmente e puntualmente utilizzati andando ad integrare ed arricchire il dato degli usuali parametri fisico-chimici.
I macroinvertebrati acquatici assumono quindi un ruolo importantissimo in quanto bioindicatori in un ecosistema molto fragile come quello dei corsi d’acqua montani. Essere in grado di conoscere e comprendere queste comunità animali porta alla possibilità di interpretare determinati segnali e variazioni, quasi sempre di tipo peggiorativo-degenerativo, per poi tradurli in una scala di qualità di salute dell’ambiente fornendoci infine gli strumenti per una più intelligente, consapevole ed accorta gestione ambientale. Tra questi macroinvertebrati gli Hydroadephaga sono stati riconosciuti da diversi autori come “Ecological Indicators” (Caro & O’doherty, 1999; Brandmayr & Pizzolotto, 2015) o “Population Indicators”. Trovandosi all’apice della catena alimentare relativa agli invertebrati e allo specifico habitat in oggetto, la loro presenza come la relativa consistenza numerica di esemplari, di generi e di singole specie, può essere considerata un buon indicatore dello stato di salute di un dato complesso biocenotico ed indirettamente della qualità delle acque. È noto infatti come lo stato di salute delle comunità di predatori di un ecosistema sia un affidabile indicatore, a cascata verso il basso della catena trofica, della salute generale di quel dato habitat.
Nonostante l’attestata importanza dei macroinvertebrati acquatici, gli studi ecologici condotti in Sardegna su tali biocenosi si presentano piuttosto scarsi, così come emerge una certa lacuna nella conoscenza della entomofauna dulciacquicola presente in numerose aree geografiche isolane. A tal proposito, si è quindi deciso di divulgare i dati di rilevamento raccolti dal 2001 al 2016 nel territorio montano di Dolianova (Provincia di Cagliari, Sardegna Orientale), con l’intento di integrare le conoscenze locali sull’entomofauna dulciacquicola, in vista di futuri lavori ecologici che possano quantitativamente e qualitativamente valutare lo stato di salute del territorio.
AREA DI STUDIO
Inquadramento geografico
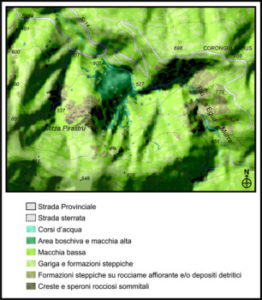
L’area di studio è localizzata nella Sardegna meridionale e ricade all’interno del Comune di Dolianova, esteso per 84,60 kmq e posto a circa 25 Km (dir.ne NE) dalla città di Cagliari (Fig. 1). Il territorio del Comune ricade nei Fogli 548 Sezioni II e III e 557 sezioni I e IV della base topografica IGM in scala 1:25.000. e nella sub-regione del Parteolla posta tra la Trexenta a NO e a O, il Siurgus a N, il Campidano di Cagliari a SO ed il complesso montuoso del Sarrabus-Gerrei da E a SE.
Estesa in una zona di transizione tra le montagne del Gerrei e la pianura del Campidano, l’area si presenta con una morfologia varia, presentando rilievi montuosi, anche con decisa acclività, che digradano in colline mioceniche dai versanti regolari e infine nelle pianure sottostanti. L’altitudine decresce procedendo verso Sud-Ovest, le cime più elevate sono situate sullo spartiacque orientale ed hanno quote comprese tra i 630 metri nel Monte Santu Miali (dove nasce il Rio Ciandara-San Giovanni) ed i 932,90 metri del Monte Tronu (dove nasce il Riu Maidana-Flumineddu). Le aree pianeggianti si estendono verso Est fino alle falde del complesso montuoso del Gerrei e si inseriscono tra i rilievi per diversi chilometri attraverso le valli dei Rii Maidana, Maidana Mannu e Bardella.
Idrografia
I corsi d’acqua presenti in questa zona fanno parte del bacino idrografico del Riu Mannu che a sua volta confluisce nel Riu Flumini Mannu per sfociare infine nello Stagno di Cagliari (S. Gilla) e di lì a mare nel golfo di Cagliari.
La rete idrografica si presenta molto ramificata in piccoli corsi d’acqua che, quasi sempre, vengono attivati solo in occasione di eventi meteorici di una certa entità (e/o costanza) risultando quindi attivi per lo più dal periodo tardo autunnale sino alla tarda primavera se le precipitazioni lo permettono. Come noto infatti, il carattere principale dei corsi d’acqua isolani è la scarsa costanza della portata dovuta sia alla irregolare alternanza delle precipitazioni che alla prevalenza di rocce impermeabili su quelle semipermeabili e permeabili che impediscono un omogeneo rapporto tra l’assorbimento delle acque meteoriche e la cessione delle stesse nelle sorgenti e nei corsi d’acqua. Le sorgenti sono numerose nella stagione invernale-primaverile per poi ridursi fino a scomparire, nella maggior parte dei casi, durante l’estate.
Una grande importanza hanno avuto nella alterazione idrogeologica dei corsi d’acqua della Sardegna le opere di deforestazione operate nell’arco degli ultimi secoli, ma iniziate in tempi ben più antichi e protrattasi, in alcuni casi, fino agli anni 50 e 60. Più recenti, ma altrettanto dannosi in quanto insistenti su complessi cenotici già frammentati, indeboliti e vulnerabili, i danni causati dagli incendi di natura antropica.
Il contrarsi delle aree boschive (foresta primaria e secondaria) e della cosiddetta macchia alta ha avuto e continua ad avere una grossa parte nella riduzione complessiva delle precipitazioni e nella successiva opera di cessione e regolazione delle acque di origine meteorica.
Clima
Tipico delle zone mediterranee insulari con il mare che svolge una funzione di termoregolazione non limitata esclusivamente agli ambiti costieri e favorisce apporti, soprattutto notturni, di umidità in particolar modo in presenza di perturbazioni e correnti provenienti dai quadranti meridionali. La “corona” di rilievi presente da NE a SE dell’area in questione favorisce, in special modo in presenza di perturbazioni occidentali atlantiche frequenti nel periodo autunnale-primaverile, l’accumulo degli ammassi nuvolosi con conseguenti fenomeni meteorici, periodicamente (da XII a II) anche in forma nevosa sui rilievi (500-900 m) ed occasionalmente a quote più basse (200 m). I valori delle precipitazioni aumentano con l’altimetria, i valori relativi alla temperatura invece diminuiscono, come noto, mediamente di circa 0,65 C° ogni 100 m. La temperatura media annua per la fascia mediomontana (da 400 a 700 m) è attorno ai 13 – 14°C, le precipitazioni per la stessa fascia si attestano tra i 700 ed i 900 mm di media annua.
Clima tardo invernale e primaverile caratterizzato da temperature generalmente miti, le perturbazioni atlantiche con i loro relativamente ingenti apporti idrici innescano fenomeni di instabilità dell’aria che, anche per diversi giorni dopo il loro passaggio, danno luogo a fenomeni di nefogenesi di tipo convettivo (cumuli e cumulonembi) sui rilievi.
Clima estivo caldo, stabile e secco con le saltuarie ed occasionali eccezioni date dal formarsi (sempre sui rilevi del Sarrabus-Gerrei) di nubi di tipo convettivo e/o temporalesco (classici temporali estivi). Anche d’estate il clima è comunque generalmente meno caldo e secco della sottostante pianura del campidano, questa differenza è tanto più percepibile quanto maggiormente si sale di quota. Il clima autunnale è molto vario e si può passare da un anno all’altro da una semplice prosecuzione (allungamento) “in minus” del clima estivo per passare ad annate fresche e maggiormente piovose.
I venti dominanti sono quelli provenienti dai quadranti occidentali (Ponente e Maestrale) ed in misura minore, anche per intensità, quelli dagli altri quadranti (Grecale NE e Libeccio SO).
Inquadramento botanico
La vegetazione presente nell’area di studio è rappresentata dalle formazioni a leccete del piano meso e termomediterraneo dove predomina come specie guida dominante il leccio (Quercus ilex L.) accompagnato da altre entità significative come: Phillyrea acutifolia L., Crataegus monogyna Jacq. e Olea europaea L. var sylvestris. Le formazioni arboree di Quercus ilex L. annoverano un gran numero di esemplari di grande vetustà e dimensione, alcune in perfetta salute, altre con evidenti segni di deperimento o danni da agenti atmosferici (forte vento, fulmini) fino ad esemplari caduti al suolo del tutto o in parte (branche, rami) ed in vario stato di decomposizione. In molti esemplari danneggiati sono evidenti i segni di ricrescita e/o ripresa. Lungo i fiumi e nelle vicinanze di sorgenti crescono rigogliosi isolati esemplari di Ficus carica L., Salix alba L., Nerium oleander L., Rubus ulmifolius Schott. e Clematis vitalba L.
Nelle radure circostanti e ai margini del bosco sono presenti Cistus salviifolius L., Pistacia lentiscus L., Ferula communis L., Asparagus acutifolius L., Rosa canina L., Urtica dioica dioica L., Asphodelus macrocarpus Parl., Cynara cardunculus var. sylvestris Lam., Cichorium intybus L.. Sugli speroni, sulle creste rocciose e nei canaloni detritici è presente la tipica flora rupestre con formazioni inquadrabili in Asplenietea trichomanis (Br.-Bl. In Meier et Br.-Bl- 1934) Oberdorfer 1977.
Quando la stagione e le condizioni climatiche lo permettono, si rinvengono inoltre numerose specie fungine, sia al suolo, nel suolo, che sul legno marcescente a volte direttamente sui tronchi ancora in piedi ma più usualmente sui detriti e sui rami caduti ed in via di disfacimento. I muschi prosperano sul terreno, in prossimità dei corsi d’acqua e sulle rocce umide cosi come alla base dei grandi alberi di Leccio e/o sulle loro branche, sugli alberi morti stesi al suolo e sui relativi detriti legnosi.

MATERIALI E METODI
Lo studio degli Hydroadephaga del territorio montano di Dolianova si è svolto dal 2001 al 2016.
Sono state indagate tutte le differenti tipologie di ambienti alle varie quote altitudinali (dai 400 m sino agli 800 m) del Riu S’Egua Murra e Mitza Pirastru (sotto il Bruncu S’Alamu), quota compresa tra 500 m e 800 m s.l.m., esposizione versante N, NE, e del Riu Giorgi Porru (Fig. 2), situato più a nord. L’indagine ha interessato sia le zone dove l’acqua era più limpida e corrente (acque lotiche) che quelle che nei periodi siccitosi o in estate divengono più torbide e stagnanti (acque lentiche).
Per la cattura degli esemplari si è proceduto con le usuali tecniche che hanno visto l’utilizzo del retino per acquatici per le pozze più profonde e del comune colino per quelle con acqua bassa.
Si è poi proceduto alla sistemazione di trappole luminose per la cattura delle specie ad abitudini fototropiche che nei periodi di maggiore siccità si spostano in volo da una pozza all’altra o da un ruscello all’altro.
Tutto il materiale citato nel presente contributo è stato personalmente visionato e determinato dagli autori. Per la realizzazione di questo contributo si è rivelata indispensabile la consultazione dei seguenti testi: Fadda & Staffa, (2014); Iiriti, (2006); Ruffo & Stoch, (2007); Vigna Taglianti, (1993); Bedel & Peyerimhoff, (1925); Franciscolo, (1979); Guignot, (1931–1933); Löbl & Smetana, (2003).
Acronimi
CDC = coll. D. Cillo, Cagliari; CEB = coll. E. Bazzato, Quartu Sant’Elena; CCA = coll. C. Ancona, Ussana; CM = coll. J. Matějíček, Formánkova, Czech Republic.
RISULTATI
Si riportano i dati di cattura relativi al rilevamento di 90 esemplari di 9 specie, appartenenti a 7 generi e due famiglie.
Materiale esaminato
Dytiscidae
Stictonectes optatus (Seidlitz, 1887)
Sardegna, Cagliari, Dolianova, Riu Giorgi Porru, 30.IV.2015, 8 es., leg. Jan Matějíček (CM).
Deronectes moestus (Fairmaire, 1858)
Sardegna, Cagliari, Dolianova, Riu Giorgi Porru 30.IV.2015, 1 es., leg. Jan Matějíček (CM).
Agabus (Gaurodytes) bipustulatus (Linné, 1767) (Fig. 3)
Sardegna, Cagliari, Dolianova, Riu Giorgi Porru, 30.IV.2015, 11 es., leg. Jan Matějíček (CM). Dolianova, Riu S’Egua Murra, 1.V. 2001 5 es., leg. D. Cillo (CDC). Dolianova, Riu S’Egua Murra, VII. 2004 11 es., leg. D. Cillo (CDC).
Agabus (Gaurodytes) nebulosus (Forster, 1771) (Fig. 3)
Sardegna, Cagliari, Dolianova, Riu Giorgi Porru, 30.IV.2015, 17 es., leg. Jan Matějíček (CM). Dolianova, Riu S’Egua Murra, VII. 2004 3 es., leg. D. Cillo (CDC). Dolianova, Riu S’Egua Murra, V 2010 1 es., leg. D. Cillo (CDC).
Agabus (Gabinectes) didymus (Olivier, 1795) (Fig. 3)
Sardegna, Cagliari, Dolianova, Riu Giorgi Porru, 30.IV.2015, 5 es., leg. Jan Matějíček (CM).

Colymbetes fuscus (Linné, 1758)
Sardegna, Cagliari, Dolianova, Riu S’Egua Murra, VII. 2004 3 es., leg. D. Cillo (CDC).
Meladema coriacea Castelnau, 1834
Sardegna, Cagliari, Dolianova, Riu S’Egua Murra, VII. 2004, 13 es., leg. D. Cillo (CDC). Dolianova, Riu S’Egua Murra, 5.II. 2002, 3 es., leg. D. Cillo (CDC). Dolianova, Riu S’Egua Murra, VII. 2008 3 es., leg. D. Cillo (CDC).
Dytiscus pisanus Castelnau, 1834 (Fig. 4)
Sardegna, Cagliari, Dolianova, Riu S’Egua Murra, VII. 2008 1 es., leg. D. Cillo (CDC).

Haliplidae
Haliplus (Neohaliplus) lineaticollis (Marsham, 1802)
Sardegna, Cagliari, Dolianova, Riu Giorgi Porru, 30.IV.2015, 5 es., leg. Jan Matějíček (CM).
DISCUSSIONI E CONCLUSIONI
I dati rilevati evidenziano come soltanto tre delle nove specie riscontrate avessero un fototropismo positivo (Dytiscus pisanus Castelnau, 1834 (fig. 4), Meladema coriacea Castelnau, 1834 e Colymbetes fuscus (Linné, 1758)), mentre le restanti (Agabus (Gabinectes) didymus (Olivier, 1795), Agabus (Gaurodytes) nebulosus (Forster, 1771), Agabus (Gaurodytes) bipustulatus (Linné, 1767), Deronectes moestus (Fairmaire, 1858), Stictonectes optatus (Seidlitz, 1887) e l’Haliplidae Haliplus (Neohaliplus) lineaticollis (Marsham, 1802)) siano state catturate esclusivamente a vista mediante utilizzo di retino per acquatici o dragando il fondo con varie tipologie di retini e colini a maglie fini.
Nonostante il numero di specie rilevato non sia estremamente elevato rispetto al numero di entità presenti a livello regionale per questo gruppo, l’abbondanza di queste nei rii montani di Dolianova, attesta come il territorio sia in un buono stato di salute generale, evidenziato anche dalla presenza di molte altre abbondanti specie dulcacquatiche appartenenti a svariati ordini: Calopteryx haemorrhoidalis (Van der Linden, 1825) (Odonata, Calopterygidae) entità citate nella Lista Rossa delle Libellule italiane seppur come entità rispettivamente a Minor Preoccupazione (LC) (Riservato et al., 2014); Boyeria irene (Fonscolombe, 1838) e Aeshna sp. (Odonata, Aeshnidae), Gerris sp., Velia sp., Notonecta sp. e Nepa sardiniensis Hungerford (Hemiptera Heteroptera).
I dati di raccolta seppur ottenuti con ricerche sporadiche, ma protratte negli anni, nel territorio montano di Dolianova, rappresentano quindi una preliminare base conoscitiva, indispensabile allo svolgimento di qualsiasi successivo studio biotico ed ecologico.
RINGRAZIAMENTI
Ringraziamo tutti gli amici e colleghi che ci hanno accompagnato e hanno collaborato alle ricerche, tra questi in particolare modo Adriana Lecis, Giuseppe Cuboni, Antonio Spiga e Daniele Sechi.
BIBLIOGRAFIA
BEDEL, L. & PEYERIMHOFF, P., 1925. Catalogue raisonné des Coléoptères du Nord de 1′ Afrique (Maroc, Algérie, Tunisie et Tripolitaine) avec notes sur la faune des Iles Canaries et de Madère, première partie. – Société Entomologique de France, Paris: 321-402 [by Peyerimhoff revised new edition of BEDEL’S original work from 1885].
BRANDMAYR, P. & PIZZOLOTTO, R., 2015. Gli Insetti Come Bioindicatori. Atti Accademia Nazionale Italiana di Entomologia. Anno LXII, 2014: 63-80.
CARO, T.M. & O’DOHERTY, G., 1999. On the use of surrogate species in conservation biology. Conserv. Biol., 13: 805-814.
FADDA, A.F. & STAFFA, F., 2014. Studio di compatibilità idraulica, Geologica e Geotecnica del territorio per l’adeguamento del redigendo PUC al PAI. Comune di Dolianova, Provincia di Cagliari.
FRANCISCOLO, M.E., 1979. Coleoptera Haliplidae, Hygrobiidae, Gyrinidae, Dytiscidae. Fauna d’Italia 14: 1-804.
GUIGNOT, F., 1931–1933. Les hydrocanthares de France. Hygrobiidae, Haliplidae, Dytiscidae et Gyrinidae de la France continentale avec notes sur les espèces de la Corse et de l’Afrique du Nord française. Les Frères Douladoure, Toulouse, 1057 pp.
KOLKWITZ, R. & MARSSON, M., 1909. Ökologie der tierischen Saprobien. Beiträge zur Lehre von der biologischen Gewässerbeurteilung. Internationale Revue der gesamten Hydrobiologie und Hydrographie, 2: 126–152.
IIRITI, G., 2006. Flora e paesaggio vegetale del Sarrabus Gerrei (Sardegna sud orientale), Università degli studi di Cagliari, Dottorato di Ricerca in Botanica Ambientale ed Applicata.
LÖBL, I. & SMETANA, A., (eds), 2003. Catalogue of Palaearctic Coleoptera. Vol. 1. Archostemata Myxophga – Adephaga. Apollo Books, Stenstrup, 819 pp.
RISERVATO, E., FABBRI, R., FESTI, A., GRIECO, C., HARDERSEN, S., LANDI, F., UTZERI, C., RONDININI, C., BATTISTONI, A., TEOFILI, C. (compilatori) 2014. Lista Rossa IUCN delle libellule Italiane. Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare,Roma
RUFFO, S. & STOCH, F. (eds), 2007. Checklist and distribution of the Italian fauna. Memorie del Museo civico di Storia naturale di Verona, 2.Serie, Sezione Scienze della Vita, 17 (2006): 1–304 + CD-ROM.
VIGNA TAGLIANTI, A., AUDISIO, P.A., BELFIORE, C., BIONDI, M., BOLOGNA, M.A., CARPANETO, G.M., DE BIASE, A., DE FELICI, S., PIATTELLA, E., RACHELI, T., ZAPPAROLI, M. & ZOIA, S., 1993. Riflessioni di gruppo sui corotipi fondamentali della fauna W-paleartica ed in particolare italiana. Biogeographia, Lavori della Società italiana di Biogeografia, 16 (1992): 159–179.
AUTORI
Davide Cillo
Via Zeffiro 8, 09126, Cagliari (CA), Italy. Email: davide.cillo@hotmail.it
Cesare Ancona
Via Mascagni 3, 09020, Ussana (CA), Italy. Email: c.ancona@yahoo.it
Luisa Dessì
Via Belvedere 14, Arbus (CA) Italy. Email: flipposters66@gmail.com
Jan Matějíček
Formankova 436, 50011 Hradec Králové (CZ), Czech republik. Email: honzama@tiscali.cz
Erika Bazzato
Dipartimento di Scienze della Vita e dell’Ambiente Sezione Botanica, Università degli Studi di Cagliari Viale Sant’Ignazio da Laconi 13, 09123, Cagliari (CA), Italy. Email: erika.bazzato@hotmail.it
PDF al seguente link
Contributo_Hydroadephaga_Bruncu_Salamu
")

. Specie erbacee")
")